
Page 79 - 2023年第54卷第2期
P. 79
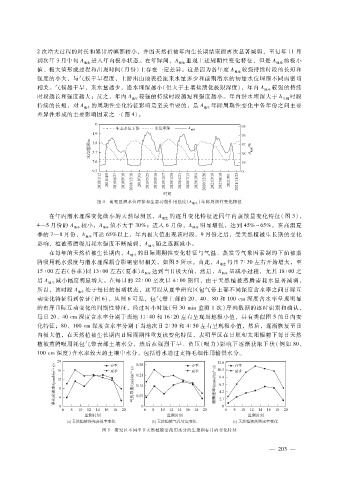
2次增大过程的时长和累计增幅都较小,并因天然植被年内生长期结束而再次显著减弱,至每年 11月
到次年 3月中旬 A 进入年内极小状态。在年际间,A 重现上述周期性变化特征,但是 A 的极小
地生 地生 地生
值、极大值形成过程和出现时间(月份)上存在一定差异。这是因为各年度 A 较强持续时段的长短和
地生
强度的小大,与气候干旱程度、上游出山地表径流来水量多少和前期潜水的初始水位埋深不同而密切
相关。气候越干旱,来水量越少,潜水埋深越小(但大于土壤盐渍化极限深度),年内 A 较强的持续
地生
时段越长和强度越大;反之,年内 A 较强的持续时段越短和强度越小。年内潜水埋深大于 h 时段
地生
生阈
持续的长短,对 A 的周期性变化特征影响是至关重要的,是 A 年际周期性变化中各年份之间主要
地生
地生
差异性形成的主要影响因素之一(图 4)。
图 4 研究区潜水位埋深和生态功能作用强度(A 地生 )年际周期性变化特征
在年内潜水埋深变化微小的天然绿洲区,A 地生 的逐月变化特征近同年内蒸散量变化特征(图 3),
4—5月份的 A 较小,A 值不大于 30%;进入 6月份,A 地生 明显增强,达到 45%~65%。在高温夏
地生
地生
季的 7—8月份,A 可达 65%以上,年内极大值出现该时段。9月份之后,受天然植被生长期的变化
地生
影响,植被蒸腾吸用耗水强度不断减弱,A 随之逐渐减小。
地生
在每年的天然植被生长期内,A 的日际周期性变化特征与气温、蒸发等气象因素制约下植被蒸
地生
腾吸用耗水强度与潜水埋深耦合影响密切相关,如图 5所示。由此,A 每日 7∶30左右开始增大,至
地生
15∶00左右(春季)同 13∶00左右(夏季)A 达到当日极大值,然后,A 呈减小过程,尤其 18∶00之
地生
地生
后 A 减小幅度明显增大。在每日的 22∶00至次日 4∶00期间,由于天然植被蒸腾需耗水显著减弱,
地生
所以,该时段 A 处于每日的极弱状态,这可以从夏季研究区包气带上部不同深度含水率之间日际互
地生
动变化特征得到佐证(图 6)。从图 6可见,包气带上部的 20、40、80和 100cm深度含水率呈现明显
的有序日际互动变化的周期性特征,经过对小时级(每 30min监测 1次)序列数据的逐时识别和确认,
每日 20、40cm深度含水率分别于当地 11∶40和 16∶20左右呈现当地极小值,具有类似图 5的日内变
化特征;80、100cm深度含水率分别于当地次日 2∶30和 4∶50左右呈现极小值,然后,逐渐恢复至日
内极大值,在天然植被生长期内日际周期性重复该变化特征,表明旱区在日照和太阳辐射下每日天然
植被蒸腾吸用耗包气带表部土壤水分,然后在强烈干旱、负压(吸力)影响下逐渐获取下伏(例如 80、
100cm深度)含水率较大的土壤中水分,包括潜水通过支持毛细作用输供水分。
图 5 研究区不同季节天然植被需耗用水分的生理指标日内变化特征
— 2 0 3 —